活動
2020年6月4日
本領域の計画研究A02中島班・分担研究者の今村 拓也 博士が、広島大学大学院 統合生命科学研究科 教授に就任しました
個性創発研究を基礎に新たなフィールドへ
今村 拓也(広島大学大学院統合生命科学研究科)
新しい研究環境について
本年4月に、7年間お世話になった九州大学から異動し、広島大学大学院統合生命科学研究科生命医科学プログラムに着任致しました。統合生命科学研究科(https://www.hiroshima-u.ac.jp/ilife)は1専攻とし、体系的な教育カリキュラムを提供する7つの基盤的な学位プログラムで構成されます。従来、理学部・生物生産学部・工学部・総合科学部および関連大学院に配置されていた各生命科学研究分野を統合し、教育効果及び現代社会のニーズの観点から分けてあります。この中で、生命医科学プログラムは新規に設置されたプログラムとして融合的な役割が期待されています。また、統合生命科学研究科は、卓越大学院プログラム(ゲノム編集先端人材育成プログラム:https://genome.hiroshima-u.ac.jp)など、いくつかの教育研究拠点コアプログラムも運営しながら進めています。
講座としては情報生理学研究室を担当することになりました。当研究室における研究テーマは多岐にわたり、ホヤ・アメフラシ・イボニシ・マウス・サル・ヒトを対象に、ゲノムから細胞・個体そして生態系にいたるまで、複数の階層を結ぶメカニズムを解く研究を推進しています。ダーウィン生誕から200年を超え、生物多様性学はポストゲノム時代に連動して分子進化から個体・集団進化に切り込む段階に入るべき時期に来たのだと思います。私のプロジェクトは下述しますが、「タンパクになれないノンコーディングRNA(ncRNA)によるエピゲノム形成とその種間多様性」です。研究対象生物の異なるメンバー達と、進化とは如何に起こるのか、を常に意識した議論をしつつ、脳の多様化・高度化戦略とその分子基盤の理解にも結びつけて行きたいと夢見ています。
ラボを立ち上げる経験
最初に教員として赴任した基礎生物学研究所には、客員部門という名称で5年を期限とする研究室が存在しました。東京大学大学院農学生命科学研究科の森裕司先生を客員教授、名古屋大学生命農学研究科の束村博子先生を客員准教授とする行動生物学研究部門が2003年度に始まることになり、私は専任の助手(のちに助教)として着任・研究を開始しました。常駐するスタッフは私ひとりでしたので、研究環境の立ち上げには随分と苦心したことを思い出します。同時に研究費獲得が全て自分自身にのしかかるというプレッシャーは相当なものでした。勿論、揃えられない機器がたくさんありましたので、逆に躊躇なく近くの研究室に出向いて使いたいものをどんどん使わせてもらおうという立場で活動しました。特に、長濱嘉孝先生や諸橋憲一郎先生(現・九州大学医学研究院・教授)の研究室を中心に出向いて、支障なく研究に打ち込めたことは大きな財産です。今思えばこのときの経験がその後の3箇所での研究環境の立ち上げに大きく役立っているように思います。諸橋先生には特定領域「性分化機構の解明」の公募研究に私の研究が採択された後に声を掛けていただき、2ヶ月に一度のプログレスレポートに参加しながらラボ運営を見せていただけたのも充実した経験でした。プログレスレポートはラボにより異なる方式があると思いますが、一週間のスパンで1-2時間を割き担当を持ち回るケースが多いように思います。それに対して、2日まるまる実験を止めて全員がプレゼンする形式です。2ヶ月という単位は絶妙だったように感じました。一週間の中で研究時間を止めなくて済みます。データがとれなかったという言い訳はききません。みんなの研究の進展具合が一目瞭然です。今のラボでも真似させていただいております。
次の赴任地は京都大学のグローバルCOEプログラム「生物の多様性と進化研究のための拠点形成」でした。プログラムリーダーの阿形清和先生(現・基礎生物学研究所・所長)には大変お世話になりました。このプログラムは、端的に言えば、学生と研究の解放区を作る、というのが目標で、阿形先生の強力な後押しもありましたおかげで、グローバルCOE准教授として最大6名のメンバーとともに、ncRNA研究を進めることができました。このとき、いわゆる次世代シーケンサーの利用とそれに伴うバイオインフォマティクスを学生たちとともに一から習得できたのが私にとっては非常に大きい出来事でした。バイオインフォマティクスときくと、実験研究者にはなかなかハードルが高く、でもできるようになったら凄いのだろうな、と漠然と考える方が多いかもしれません。実際、私もそうでした。この点、グローバルCOEには西村理さん(現・理化学研究所)という実験研究にもバイオインフォマティクスにも精通したスタッフがおられましたので、恥を忍んで教えを乞い、それこそUNIXのコマンドの基礎からワークステーションの立ち上げまで、何でも懇切丁寧に教えていただきました。やはりやってみなければわからないもので、やってみると面白いものです。こんなことできるのではないか、と思いついたら、まずは「グーグル先生」に問い合わせて、大体なんでも無料のプログラムや実行コマンドの情報に行きあたれるものです。自分なりにカスタマイズして、また、学生たちとその技を教え合いながら、pancRNAと名付けた一連のプロモーターncRNAデータベースを仕上げることに成功し、そこから一般則(pancRNAは下流遺伝子を特異的に活性化できる)を見出すことができました。
スタッフとして沢山のラボを経験できたというのは私にとっての財産です。前任地の九州大学では、中島欽一先生に大変よくしていただきました。中島先生の凄いところは、まず勉強量です。教授が一番論文を読んでいるラボでした。また、神経発生分野に留まらず、国内外の沢山の研究者と知り合いで、その縁で沢山の研究者ネットワークに参加することができるようになりました。本新学術領域研究「多様な『個性』を創発する脳システムの統合的理解」に参画することができたのも、そのネットワークの一つです。大隅典子先生を代表とする研究班の中で、さらに沢山のメンバーと知りあい、沢山の情報交換をすることができました。このように書いてみると、若手・中堅研究者にとって、文部科学省研究費ベースの集まりというのは、時間限定の中で、それぞれの個性ある研究の進展を互いに見ながら、負けないぞと切磋琢磨していくことができるところに醍醐味があるのでしょう。
私のプロジェクトについて
脳の形態学的・機能的な違いは遺伝的に98%の相同性を示すヒト・サルでも明らかであり、実験動物として汎用されるマウスも、殆どの遺伝子セットを共通に利用していながら、独特な神経系を獲得しています。一方、ncRNAセットは種間多様度が極めて高いです。私は、ほ乳類脳の種特異性成立メカニズムにおいては、種特異的なncRNAが機能することを想定しています。
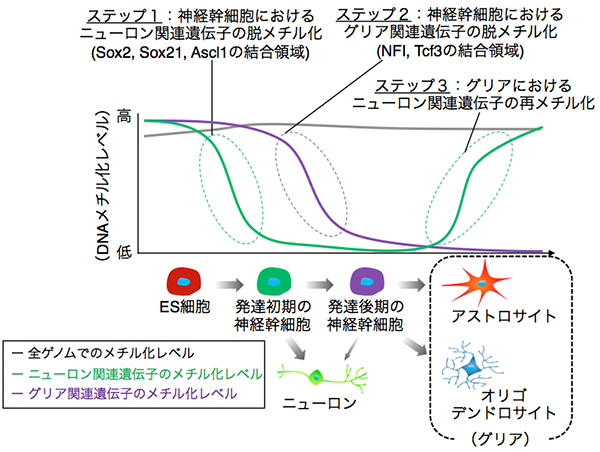
本新学術領域研究のサポートのもと得られた成果として、発達期のげっ歯類大脳神経幹細胞において、特定の転写因子をベースとした配列特異的エピゲノム変換が秩序だった脳形態形成を保証していることを明らかにしました (Sanosaka et al., Cell Reports, 2017)。エピゲノム変換のヴァリエーションが個性創発と関係している可能性があります。大脳神経幹細胞の場合、配列特異的エピゲノム変換ステップは以下の3つから成り立っています(九州大学プレスリリース, 2017 https://www.kyushu-u.ac.jp/ja/researches/view/171 :上図): I. 多能性幹細胞から誘導された神経幹細胞からは、先ずニューロンのみが優先的に産生されるが、このとき、ゲノムDNA における Sox2・Sox21・Ascl1の結合とDNA 脱メチル化が強く結びついて起こる。II. 次に、神経幹細胞は、Nuclear factor I (NFI)による DNA 脱メチル化の誘導により、ゲノム全体にわたる制御を一新し、神経幹細胞が産み出す細胞を、アストロサイトやオリゴデンドロサイトを生み出せる状態に移行させる。III. 一旦アストロサイトやオリゴデンドロサイトへと分化した細胞では、ニューロンで機能する遺伝子発現制御領域を再メチル化し強制的に OFF にすることで、アストロサイトやオリゴデンドロサイトから別の細胞、すなわちニューロンへと再分化されないように固定される。
種差の観点からみたとき、例えば、霊長類ではげっ歯類の数十倍の平均寿命があることから、上記に発見した神経幹細胞の性質変化プロセスも適応的に変化しているはずであると考えています。実際に、例えば、霊長類神経幹細胞の場合、げっ歯類に比べて、アストロサイト分化能を獲得するのに非常に時間がかかります(4日→200日)。ヒト神経幹細胞モデル系では、酸素分圧の改変によりアストロサイト分化を促進できることからも、個性創発メカニズムと同様に、遺伝的素因に加えてエピゲノム変換の種差が神経系細胞の形質発現と強く結びついていることが考えられます。ニューロンについても、一生を通じて長く生きる必要があるため、動物種の違いに基づいた環境変化を受容できるように、種にしたがった別々の強固なクロマチン基盤を備えている可能性が高いと考えています。
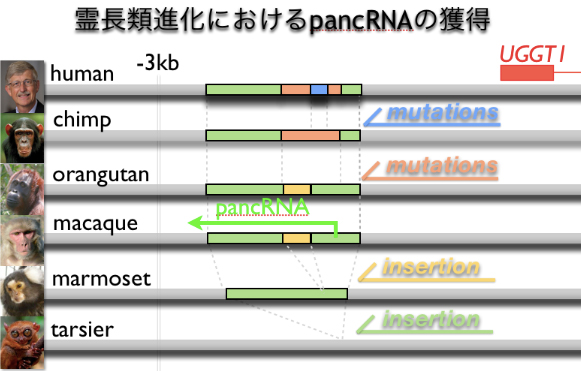
実際に最近、 5種5組織の計25サンプルの大規模比較トランスクリプトーム解析を行い、種特異的プロモーターncRNA(pancRNA)を進化的に獲得すると、特定の細胞群でのみ遺伝子発現を達成できることを発見しました(Uesaka et al., BMC Genomics, 2017:上図)。pancRNAは下流遺伝子のエピジェネティクスを活性型に変換し下流遺伝子を特異的に発現上昇させることのできるncRNAです。さらに、最近になって実際に、種特異的pancRNAを神経幹細胞にて操作すると、増殖と分化の傾向を改変できることを見出しつつあります。したがって、種を超えて保存されていない配列の獲得が、種特異的pancRNA産生/エピジェネティック変換を介して、遺伝子発現スイッチの獲得に至ると考えています。
現在、マウスES細胞や霊長類iPS細胞を活用しながら、CRISPR/CAS9システムなどを用いた種特異的pancRNA導入技術を基礎とした研究を進め、神経活動のネットワーク化改変にチャレンジしています。特に、異動物種の形質をマウス脳オルガノイドに賦与する技術の開発とバイオインフォマティクスを駆使した分子メカニズムの網羅的解明に挑もうとしています。
おわりに
「ncRNA」、「エピゲノム」、「進化」、「霊長類脳」をキーワードに、多様な人材を受け入れ、活発な分野間交流を通して私たちも進化していきます。研究室訪問は随時受け付けております(ただ、当分はオンラインにてということになりそうです:現メンバーとの写真参照)。大学院生、研究生として参加を希望する方、私たちの研究に興味を持たれた方は個別にお気軽にご連絡ください。参考URLは以下です。

今村拓也:緊急事態宣言前の3月に尾道にて
5月のリモートミーティングの様子

亀田 朋典 さん

安 博洋 さん

森下 文浩 さん

植木 龍也 さん